Il miglioramento genetico per caratteri qualitativi, accanto al miglioramento genetico per produzione ha consentito all’uomo di trasformare positivamente le produzioni agricole. Per quanto riguarda i cereali, si è assistito al perfezionamento dei metodi per selezionare una serie di composti importanti dal punto di vista nutrizionale e tecnologico. In frumento e in mais l’obiettivo del miglioramento sono state le proteine, nel primo caso in funzione della qualità tecnologica del prodotto finale (pasta o pane), nel secondo in vista di un aumento del contenuto in aminoacidi essenziali, per combattere il problema della malnutrizione. La selezione applicata al riso si è invece focalizzata sul contenuto di amilosio, di composti antiossidanti e sull’aroma. Infine, la componente solubile della fibra è risultata essere interessante per la selezione di genotipi di orzo da utilizzare come alimenti funzionali.
Proteine del frumento per la qualità di pane e pasta
La qualità tecnologica di una farina di frumento (Triticum ssp.) è legata in modo particolare al contenuto proteico. A partire dagli anni ’80, l’attività di miglioramento genetico del frumento ha avuto un notevole impulso dalla individuazione di specifiche proteine correlate con la qualità del glutine. In particolare, è stato dimostrato che le glutenine, una delle frazioni proteiche del glutine, determinano le caratteristiche reologiche degli impasti (elasticità ed estensibilità), attraverso la formazione di polimeri proteici costituiti da subunità gluteniniche a basso (Low Molecular Weight Glutenin Subunits, LMW-GS) o alto peso molecolare (High Molecular Weight Glutenin Subunits, HMW-GS). La presenza di certe subunità HMW è infatti correlata alla qualità panificatoria delle farine, valutata attraverso test reologici: a ogni specifico componente delle HMW viene associato un punteggio qualitativo, come riportato in Tabella 1 (Pogna et al., 1989). La composizione in glutenine, inoltre, è un parametro determinato geneticamente e non influenzato dalle condizioni ambientali: rappresenta perciò un metodo semplice e ripetibile per selezionare le varietà da sviluppare. Anche le gliadine costituiscono una componente importante del glutine, ma a differenza delle glutenine non formano polimeri. Una forte correlazione è stata osservata tra la presenza della gliadina γ-45 e la buona qualità della pasta, in termini di tenuta alla cottura (Damidaux et al., 1978; Payne, 1987). La conseguenza di questa osservazione è stata che la quasi totalità delle varietà di frumento duro attualmente in commercio esprime la gliadina γ-45, con una significativa riduzione della variabilità genetica disponibile.
|
|
Subunità di glutenine HMW |
||
|
Score |
Glu-A1 |
Glu-B1 |
Glu-D1 |
|
6 |
|
17 + 18 |
5 + 10 |
|
5 |
2 |
7 + 9 |
|
|
4 |
|
7 + 8 |
|
|
3 |
1 |
|
|
|
2 |
|
7 |
2 + 12, 5 + 12 |
|
1 |
|
20, 6 + 8 |
4 + 12 |
Tabella 1. Score relativi alla qualità di subunità di glutenine ad alto peso molecolare
in frumento tenero sulla base del test alveografico. Da: Pogna et al., 1989.
Mais: miglioramento qualitativo delle proteine
Il contenuto proteico del mais si aggira intorno al 9-11% sul peso secco del seme. La frazione più abbondante è rappresentata dalle zeine, proteine a basso peso molecolare solubili in alcool. Esse rappresentano il 50-70% delle proteine presenti nell’endosperma di mais e sono molto ricche in alcuni aminoacidi quali glutammina, leucina e prolina, ma povere in lisina e triptofano. Nel 1920 è stato scoperto un mutante chiamato Opaco-2 (Singleton, 1939) caratterizzato da un alto contenuto di lisina. In questo mutante la frazione di α-zeine era meno abbondante del normale, mentre risultava aumentato il contenuto in altre frazioni proteiche più ricche in lisina. Le piante che portano la mutazione opaco-2 però, pur avendo caratteristiche nutrizionali favorevoli, presentano alcuni aspetti negativi: basse rese e un endosperma farinaceo e soffice, che rendeva la granella più suscettibile agli attacchi di patogeni e alle malattie. Si è cercato quindi di migliorare il genotipo opaco-2 attraverso la combinazione con uno o più geni in grado di modificare la consistenza dell’endosperma (ad esempio il gene sugary-2, Su-2). Come risultato sono state ottenute linee di mais caratterizzate da un alto contenuto di lisina insieme alla tessitura vitrea della granella. A partire da questi genotipi sono stati rilasciati a partire dal 1985 diversi ibridi chiamati QPM (quality protein maize), introdotti nei paesi del Sud del Mondo per contrastare il problema della malnutrizione (Sofi et al., 2009).
Selezione di riso per contenuto in amilosio, componenti funzionali e aroma
La molecola dell’amido è composta da amilosio e amilopectina. L’amilosio (il cui contenuto è variabile tra 7 e 33%), è considerato il componente in grado di influenzare maggiormente il comportamento del granello di riso alla cottura e alla masticazione: se la sua percentuale è elevata, migliora la capacità dell’amido di assorbire acqua durante la cottura e il granello mantiene una certa consistenza riducendo la collosità superficiale. In caso contrario, i granelli in seguito alla cottura mostrano una notevole collosità e non risultano adatti alla preparazione di risotti o minestre. Il contenuto di amilosio è controllato dal gene waxy che codifica per l’enzima amido sintasi. Questo gene presenta dei SNP (Polimorfismi a Singolo Nucleotide) che sono correlati con il contenuto di amilosio; attraverso l’uso di questi marcatori molecolari è possibile selezionare genotipi per il breeding aventi alto contenuto di amilosio.
Recentemente la selezione di genotipi di riso ad alta qualità nutrizionale si è focalizzata sulle proprietà antiossidanti. Un studio riguardante i risi pigmentati (Finocchiaro et al., 2010) ha dimostrato che i risi rossi sono ricchi di proantocianidine, i risi neri di antocianine, ed entrambi hanno una capacità antiossidante totale molto più alta rispetto ai risi bianchi. In particolare la varietà nera Artemide si è rivelata essere ricca non solo di antocianine e proantocianidine, ma anche di polifenoli.
Anche l’aspetto dell’aroma è stato un target del miglioramento genetico. Una varietà aromatica è caratterizzata dal possedere un granello che emana un particolare profumo. La 2-acetil-1-pirrolina (2AP) è una delle molecole aromatiche responsabile dell’aroma del riso ed è presente in tutti i genotipi. Tale molecola viene degradata dall’enzima betaina aldeide deidrogenasi (BAD2), che nelle varietà aromatiche ha una delezione di 8 nucleotidi e quindi perde la sua funzione (Figura 1).
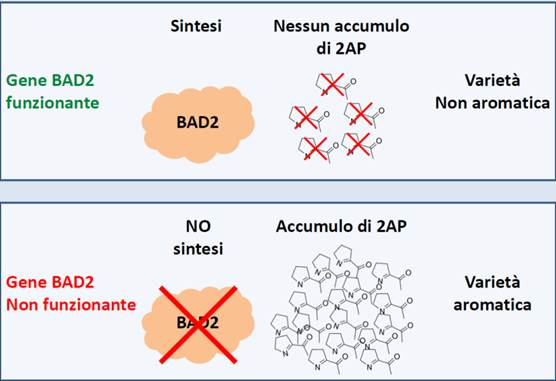
figura 1 - Differenze gene BAD2 in varietà aromatiche e non aromatiche di riso
Gli individui eterozigoti non sono aromatici, ma il carattere può essere fissato in omozigosi recessiva nelle successive generazioni. La varietà aromatica Tigre, di cui l’Ente Nazionale Risi è costitutore, è stata ottenuta tramite l’utilizzo della MAS (Marker Assisted Selection) che ha permesso di rilevare all’interno della progenie di alcuni incroci specifici finalizzati all’ottenimento di varietà aromatiche, le linee che portavano la variante allelica associata al carattere aroma.
Orzo e composti bioattivi
Nei cereali sono presenti numerosi composti bioattivi (β-glucani, tocotrienoli, folati, fruttani, fitosteroli, composti fenolici, fitati). Nell’orzo, la fibra solubile è rappresentata soprattutto dai β-glucani, che si trovano negli strati più interni della cariosside. Questi composti sono importanti per vari motivi: riducono il colesterolo plasmatico, condizionando la biosintesi di quello endogeno; permettono una selezione della flora intestinale grazie all’attività fermentativa; abbassano il livello di glucosio postprandiale e la conseguente risposta insulinica; rallentano lo svuotamento gastrico, grazie alla loro viscosità, e procurano così un senso di sazietà. Sulla base dell’importanza di questi composti, la Regione Emilia Romagna negli anni 2005-2007 ha finanziato il progetto di ricerca “Filiera dell’orzo per alimenti funzionali”. Nel corso del triennio, partendo da genotipi di orzo ad alto tenore di β-glucani, opportunamente valutati e selezionati, si è passati alla messa a punto di tecnologie in grado di arricchire ulteriormente i materiali in β-glucani, selezionando specifiche frazioni di farina; sono stati quindi prodotti e valutati pane, pasta e dolci con miscele di farine di frumento tenero/duro e orzo, che potrebbero essere inseriti in una alimentazione salutare.
collegamento consigliato
Differenze gene BAD2 in varietà aromatiche e non aromatiche di riso
Differenze gene BAD2 in varietà aromatiche e non aromatiche di riso. In: La selezione assistita da marcatori (SAM), http://www.enterisi.it/upload/enterisi/gestionedocumentale/PosterFinaleSAM2_qualit%c3%a0_granello_784_12046.pdf
Gli esempi sopra riportati illustravano alcune possibili strategie per la selezione di genotipi interessanti per caratteri qualitativi. Le selezioni svolte in passato avevano solamente mirato ai parametri agronomici, migliorando la resa e la resistenza ai fattori di stress, sia biotici che abiotici. In seguito l’attenzione della ricerca agronomica si è spostata anche verso altri aspetti, in particolare la caratterizzazione dal punto di vista nutrizionale. La tendenza più recente dei consumatori, perlomeno nei paesi occidentali, è infatti quella di guardare agli alimenti non solo come fonte di energia ma anche come uno strumento per la prevenzione di alcune patologie che appaiono essere legate a uno stile alimentare poco equilibrato.
Damidaux R., Autran J. C., Grignac P. e Feillet P. (1978). Mise en évidence de relations applicable en sélection entre l’electrophoregramme des gliadines et les propriétés viscoélastique du gluten de Tr. durum Desf. C.R. Acad. Sci. Paris Serie D: 701-704.
Finocchiaro F., Ferrari B, Gianinetti A. (2010). A study of biodiversity of flavonoid content in the rice caryopsis evidencing simultaneous accumulation of anthocyanins and proanthocyanidins in a black-grained genotype. Journal of Cereal Science 51: 28–34.
Gianinetti A. (2008). Negli alimenti funzionali l’orzo fa la differenza. Agricoltura Giugno: 94-96.
MAS su riso per aroma e amilosio (poster)
Payne P. I. (1987). Genetics of wheat storage proteins and the effect of allelic variation on breadmaking quality. Annual Reviews Plant Physiology 38: 141-53.
Pogna N., Gazza L. (2004). Genotipi di grano duro ad elevata qualità panificatoria: risultati e prospettive. In: Ottimizzazione delle materie prime, del processo di lievitazione e del confezionamento nella produzione industriale del pane e prodotti da forno convenzionali e dietetici, Consorzio Gian Pietro Ballatore, pp. 12-30
Pogna N.E., Mellini F., Beretta A., Dal Belin Peruffo A. (1989). The high-molecular-weight glutenin subunits of common wheat cultivars grown in Italy. Journal Genetics & Breeding 43: 17-24.
Singleton R.V. (1939). Recent linkage studies in maize. Opaque endosperm-2 (o2). Genetics 24: 61
Sofi P.A, Wani S.A., Rather A.G, Wani S.H. (2009). Review article: quality protein maize (QPM): genetic manipulation for the nutritional fortification of maize. Journal of Plant Breeding and Crop Science 1(6): 244-253.

