Le piante rispondono all’attacco di patogeni attraverso due strategie: la prima coinvolge le barriere strutturali-morfologiche e alcuni composti biochimici naturalmente presenti, la seconda consiste in una risposta indotta dal patogeno, cioè una serie di metaboliti secondari prodotti solo in caso di attacco. Queste risposte sono spesso regolate a livello genetico, per cui è possibile trovare delle differenze nella risposta ai patogeni in genotipi diversi. Per questo uno dei metodi per la difesa della produzione agricola è la selezione di genotipi che abbiano caratteri di resistenza, attraverso uno screening delle piante in condizioni di attacco artificiale del patogeno, oppure attraverso l’utilizzo di specifiche tecniche di biologia molecolare (MAS). Due metodi di screening di genotipi di mais resistenti a diabrotica e al fungo F.verticillioides sono stato messi a punto.
Una strategia per difendere la produzione agricola è la selezione, tra tutti i genotipi disponibili, di piante che presentano caratteri di resistenza ai patogeni. Le piante infatti dispongono di sistemi di resistenza geneticamente regolati determinati da una combinazione di meccanismi che coinvolgono una serie di proteine e molecole organiche presenti prima o durante l’attacco di patogeni.
Meccanismi di difesa delle piante
Le prime forme di resistenza vegetali sono rappresentate da difese costitutive, cioè naturalmente presenti nelle piante (morfologiche, strutturali o biochimiche). Si tratta per esempio di cere, lignina, cellulosa o proteine della parete cellulare che costituiscono barriere strutturali. A queste si aggiungono una serie di metaboliti secondari con funzione antimicrobica, presenti in normali condizioni fisiologiche. Si tratta per esempio di terpenoidi, steroidi e glucosinolati e proteine antifungine come le saponine (Tava e Avato 2006) o le RIPs (Ribosome-Inactivating proteins), una famiglia di proteine che catalizza l’inattivazione dei ribosomi dei patogeni (Lanzanova et al. 2009).
Una seconda strategia di resistenza è rappresentata da difese indotte in risposta all’attacco patogeno. Esse sono attivate da specifici meccanismi di riconoscimento gene specifici tra molecole che fungono da recettore della cellula vegetale (le proteine R) e molecole segnale prodotte dal patogeno chiamate elicitori. Il riconoscimento e quindi l’interazione pianta-patogeno, avviene grazie al legame tra elicitori e proteine R che, tramite un cambiamento conformazionale, innescano una cascata di segnale mediata da una classe di proteine (proteine G). Questa reazione attiva diversi meccanismi di difesa tra cui la produzione di specie reattive dell’ossigeno (ROS) con ruolo antimicrobico, la morte cellulare localizzata nel punto di infezione con lo scopo di bloccare l’avanzamento del patogeno, modifiche della parete cellulare e la sintesi di proteine indotte dalla patogenesi (pathogenesis-related, PR) (Kitajima e Sato, 1999).
Come avviene la selezione
La costituzione di piante capaci di sottrarsi, in tutto o in parte, ai danni provocati dai patogeni è un obiettivo tradizionale nell’ambito del miglioramento genetico. Lo sviluppo di tecniche affidabili e idonee per lo screening e la valutazione di genotipi resistenti all’attacco di patogeni costituisce perciò un importante strumento nei programmi di breeding. Bisogna considerare però che in condizioni di scarsa incidenza della patologia (funghi, insetti, virus) risulta difficile poter mettere in evidenza un’eventuale variabilità nella risposta dei genotipi sotto studio. L’approccio utilizzato in esperimenti che hanno come obiettivo la selezione per resistenza è quindi una forzatura del sistema, che consiste nell’infettare artificialmente le piante col patogeno. L’attacco artificiale deve i) mimare le modalità di accesso del patogeno verso la pianta, ii) avvenire, dove possibile, in ambiente controllato, in modo da eliminare tutte le variabili che potrebbero influenzare la risposta, iii) essere riproducibile. La valutazione del danno dovuto all’infezione artificiale viene affiancata alla valutazione della presenza del patogeno nell’ambiente naturale, quindi sulle piante non infettate (Figura 1).
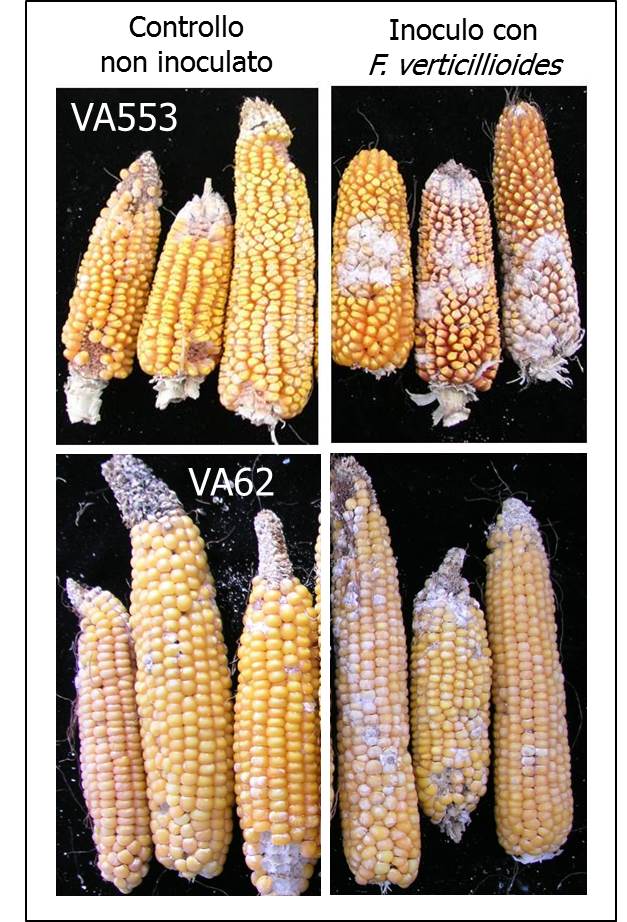
figura 1 - Spighe di mais con F.verticillioides
Accanto ai metodi di screening per le resistenze è possibile studiare anche a livello molecolare i geni che vengono attivati in condizioni di stress biotico. Un esempio di questo approccio è descritto in un recente lavoro di Lanubile et al. (2010), in cui sono stati identificati geni differenzialmente espressi in genotipi di mais di derivazione canadese con diversi livelli di resistenza in seguito a infezione artificiale da parte di Fusarium verticillioides attraverso le sete e la cariosside. Possono inoltre venire applicati anche metodi di miglioramento genetico avanzato, che consistono nell’introduzione di resistenze genetiche mediante l’uso della selezione assistita da marcatori associati al gene da introdurre (Marker Assisted Selection, MAS). Una possibile limitazione dell’utilizzo di resistenze genetiche è data dall’evoluzione dei patogeni che possono “superare” l’efficacia dei geni di resistenza utilizzati. Per superare questo problema è possibile attuare un particolare approccio di miglioramento genetico chiamato gene pyramiding, che consiste nell’accumulare in un medesimo genotipo più geni di resistenza nei confronti di un determinato patogeno. Questo processo, attuabile esclusivamente con la MAS, consente di ottenere resistenze durevoli, in quanto il patogeno per essere attivo dovrebbe acquisire la capacità di superare più geni di resistenza.
Esempi di selezione di genotipi resistenti
Un esempio di selezione attraverso metodi di screening è stata operata sul mais nell’ambito del progetto IDIAM (Interventi per contrastare la diffusione e i danni da DIAbrotica nella Maiscoltura italiana), che aveva come focus le strategie di resistenza contro l’infestazione da diabrotica. A tal fine si è reso necessario mettere a punto un protocollo in ambiente controllato (fitotrone), che ha permesso di seguire il corretto sviluppo e la vitalità delle larve di diabrotica a partire dalla schiusa delle uova mantenute in diapausa a 4 °C. La schiusa delle uova direttamente a contatto con seme di mais germinato in scatole Magenta con terriccio offre alle larve neonate la possibilità di avere subito a disposizione le radici di plantule di mais di cui nutrirsi. È stata osservata la crescita controllata in fitotrone di alcuni ibridi di mais, presenti nella rete di prove agronomiche, in presenza o meno (controllo) di infestazione artificiale. Dopo 21 giorni dalla semina è risultato evidente il danno provocato dalle larve sulle radici delle plantule di mais che, nei campioni infestati, risultano essere maggiormente ridotte rispetto ai controlli che invece non hanno subìto danno, confermando quindi l’efficacia del metodo di infestazione adottato (Figura 2). La presenza di correlazione tra il peso dell’apparato radicale di ciascun genotipo in situazione di controllo e infestato ha permesso diconfermare che gli ibridi che presentano apparati radicali più sviluppati sono in grado, anche con attacco larvale, di difendersi meglio rispetto a ibridi con apparati radicali più ridotti.

figura 2 - Piante di mais infestate da diabrotica
Un altro esempio riguardante il mais è la messa a punto di sistemi di inoculo artificiale con spore di Fusarium verticillioides (Ferrari e Balconi, 2008). Le tecniche di inoculo artificiale adottate sono le seguenti: i) tramite sete (non invasiva, spray), in cui la sospensione di spore viene vaporizzata sulle sete della spiga primaria, 4- 8 giorni dopo l’impollinazione (DAP); ii) tramite sete (invasiva, siringa), in cui la sospensione di spore viene iniettata nel canale delle sete della spiga primaria, a 4-8 DAP; iii) tramite cariossidi (invasiva), in cui la sospensione di spore viene veicolata alla spiga primaria (zona centro- basale) tramite un’incisione di tre cariossidi, a 15-20 giorni DAP. Il grado di attacco fungino viene registrato alla raccolta, tramite scala di valutazione basata sulla percentuale di cariossidi con segni visibili di infezione, assegnando un punteggio da 1 (0 cariossidi con infezione visibile) a 7 (76-100% cariossidi/spiga con infezione visibile) (Reid et al. 1996). Questi approcci hanno consentito in anni recenti di valutare e selezionare genotipi di mais resistenti al patogeno, anche in relazione al livello di micotossine prodotte dopo l’infezione (Balconi et al., 2014).
Spighe di mais con F.verticillioides
Spighe di due varietà tradizionali valutate per la resistenza al patogeno fungino Fusarium verticillioides. (Foto A. Torri)
Piante di mais infestate da diabrotica
Piante di mais infestate artificialmente da larve di diabrotica, e relativo controllo (Foto C. Lanzanova)
Nell’ambito della difesa delle colture agricole dagli organismi patogeni, la ricerca rivela il suo potenziale impatto positivo per ridurre l’utilizzo di sostanze chimiche, favorendo non solo un risparmio sui costi di coltivazione ma anche una riduzione dell’impatto ambientale. Le tecniche di ingegneria genetica non sono praticabili in questo contesto, in quanto oltre a essere vietate dalla legislazione vengono apertamente osteggiate dall’opinione pubblica. La descrizione della variabilità esistente in termini di sintesi di molecole con funzione antifungina o antimicrobica, che limita l’attacco alle piante da parte di un patogeno, la sua diffusione all’interno dei tessuti o la sintesi di molecole tossiche per gli animali e l’uomo, descrizione realizzata attraverso screening a livello di DNA o mediante prove in campo, resta quindi al momento l’unico strumento disponibile. È perciò importante proseguire e approfondire questo campo di studi, per comprendere sempre meglio il rapporto tra pianta e patogeno e trovare strategie di difesa sempre più efficaci.
Balconi C., Berardo N., Locatelli S., Lanzanova C., Torri A., Redaelli R. (2014). Evaluation of ear rot (Fusarium verticillioides) resistance and fumonisin accumulation in Italian maize inbred lines. Phytopatologia Mediterranea 53 (1): 14−26.
Ferrari A., Balconi C. (2008). Tecniche di inoculo fungino per la valutazione di genotipi di mais. Dal Seme 1: 38-40.
Ferreira R.B., Monteiro S., Freitas R., Santos C.N., Chen Z., Batista L.M, Duarte J., Borges A., Teixeira A.R. (2007). The role of plant defense proteins in fungal pathogenesis. Molecular Plant Pathology 8 (5): 677–700.
Kitajima S. and Sato F. (1999). Plant pathogenesis-related proteins: molecular mechanism of gene expression and protein function. Journal of Biochemistry 125: 1-8.
Lanubile A., Pasini L., Marocco A. (2010). Differential gene expression in kernels and silks of maize lines with contrasting levels of ear rot resistance after Fusarium verticillioides infection. Journal of Plant Physiology 167: 1398–1406.
Lanzanova C., Giuffrida M.G., Motto M., Baro C., Donn G., Hartings H., Lupotto E., Careri, Elviri L. Balconi C. (2009). The Zea mays b-32 ribosome-inactivating protein efficiently inhibits growth of Fusarium verticillioides on leaf pieces in vitro. European Journal of Plant Pathology 124: 471-482.
Miglioramento genetico e biodiversità. (2009). Dipartimento di biologia e produzione vegetale. Consiglio per la Ricerca e la sperimentazione in Agricoltura A cura di Di Fonzo N. e D’Andrea F.
Reid L.M., Hamilton R.I., Mather D.E. (1996a). Screening maize for resistance to Gibberella ear rot. Technical Bull 1996-5E, Research Branch, Agriculture and Agri-Food Canada.
Strategie innovative di difesa dalla diabrotica del mais. L’Informatore Agrario, 4/2014.
Tava A., Avato P. (2006). Chemical and biological activity of triterpene saponins from Medicago species. Natural Product Communications 1(12): 1159-1180.

